Lab 2 Plant Structure
The morphology and
anatomy of fossil plants contains a wealth of information on the function,
physiology, ecology, and life habit of ancient plants. Plant morphology can
also yield clues to taxonomic and evolutionary relationships. Consequently,
background in plant structure is a prerequisite for studying land plant
evolution. This lab reviews plant structure, especially cell and tissue types,
and the arrangement of the vascular system. We provide only the most basic
information here. For a more comprehensive review of plant anatomy and
morphology, consult the following references:
Bierhorst, D.W. 1971. Morphology of Vascular Plants. .
Esau, K. 1965. Plant
Anatomy, second edition. Wiley,
Foster, A. and E.M. Gifford. 1974. Comparative
Morphology of Vascular Plants. . Freeman,
Raven, P.H., R.F. Evert, and H. Curtis. 1981. Biology of Plants., third edition. Worth,
Basic
Organization
Compared
with animals, plants have a relatively simple design. Most land plants consist
of a stem
or axis,
which functions for support and contains the conducting tissues of the plant.
The stem usually supports light-gathering and photosynthetic structures called leaves (VG 1:1)(VG 1:2), and the
plant's reproductive structures, which may go by the names flowers,
sporangia (VG 1:3), cones, or
any number of others depending on the taxon. Land
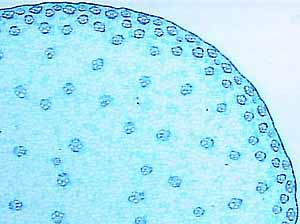
plants are anchored to their substrate by roots (VG 1:4) or rhizomes,
which are really underground stems. Although there are relatively few basic
parts to plants, each part can take on an amazing variety of forms. Compare
redwood or oak trees and the bluegrass from the surrounding lawn. Both have
stems, leaves, and reproductive structures, but they look very different.
The variety of
stem form (woody or non-woody, densly branching or
un-branched) gives plants a variety of growth forms.
For example, "tree" "bush" and "herb" are
important classes of growth forms. Plants with different growth forms often
have different life histories and ecologies. Since life history and ecology are
important features that are modified during evolution, growth form is an
important feature of plants and lineages. Some plants occupy a number of growth
forms depending on the conditions under which they live, or at different points
in their life cycle. The terms "tree", "bush" and
"herb" also have colloquial meanings that make them difficult to
define precisely in a scientific sense.
Plant Cell and Tissue Types
PARENCHYMA
Parenchyma
cells, the progenitor of all other cell types, are composed of thin walled,
globular, more or less undifferentiated cells. Parenchyma cells comprise many
soft tissues of plants (e.g., pith, cortex,
leaf mesophyll, etc.). These cells also compose the
horizontal rays in wood. Parenchyma cells retain the ability to divide
throughout their lives, so they are important in vegetative regeneration and
wound healing. For example, roots growing from a stem cutting are created and
differentiate from parenchyma cells that are scattered throughout the stem and
spring into action when cued by hormonal changes that a new structure is
needed. Most of the "work" of plants (e.g., photosynthesis,
carbohydrate storage, metabolism, secretion, and biosynthesis) occurs in
parenchyma cells. As parenchyma is incorporated into vascular tissue (rays in
wood for example), it also helps in the movement of water and solutes
throughout the plant body. Because parenchyma tissue is composed of only one
cell type, parenchyma is called a simple
tissue.
COLLENCHYMA
Collenchyma tissues are composed of prismatic cells
that are commonly elongated and can occur in long strands or cylinders. Like
parenchyma cells, collenchyma is living at maturity. Collenchyma cells have thick primary walls composed of cellulose.
(Note that you can distinguish collenchyma cells from
sclerenchyma cells because of the chemical
composition of their cell walls. Different biological stains are attracted to
either cellulose or lignin. Consequently, in the most common stain system,
cellulose stains blue or green and lignin stains reddish or pink.) Because collenchyma cell walls are not lignified, the collenchyma strands are flexible, thus ideal for structural
support and protection in growing shoots or flexible structures like leaves. Collenchyma is found near the surface of cortex
in stems and along the veins of leaves, where it provides structural support
and protection against breakage.
SCLERENCHYMA
Sclerenchyma cells have thick, lignified secondary
walls, lack cell contents at maturity, and occur throughout all plant tissues.
These features make sclerenchyma tissues hard, rigid,
and somewhat brittle. Sclerenchyma cells can occur as
aggregates within ground
tissue (sclereids or stone cells or as elongated fibers.
In this context, sclerenchyma provides mechanical
strength to stems (fibers in hemp and flax) and reproductive structures (the
texture in pear flesh, the stony shells of nuts and cherry pits).
(Note that you can
distinguish collenchyma cells from sclerenchyma cells because of the chemical composition of
their cell walls. Different biological stains are attracted to either cellulose
or lignin. Consequently, in the most common stain system, cellulose stains blue
or green and lignin stains reddish or pink.)
XYLEM
Xylem
tissue functions in both water transport and mechanical support. In
non-angiosperm tracheophytes, tracheids (Figure 1.1) serve both purposes; in most
angiosperms, the xylem contains both vessel
elements, which have a larger diameter and are specialized for water
transport, and fibers
for mechanical strength.
Xylem cells
commonly have cell walls impregnated with lignin
and reinforced with spiral or ring-like thickenings that project into the lumen of
the cell (Figure 1.2). Both features reinforce the cells for mechanical
support.
|
|
|
Figure 1.1: Xylem cell types. (A) Sclereid reinforced witrh
lignin; (B) tracheid of Woodwardia, a fern
(one-sixth of cell shown); (C) Pinus, a conifer (one-third of cell
shown); (D) fiber tracheid; (E-G) angiosperm xylem
-- (E-F) tracheids, (G) vessel member. |
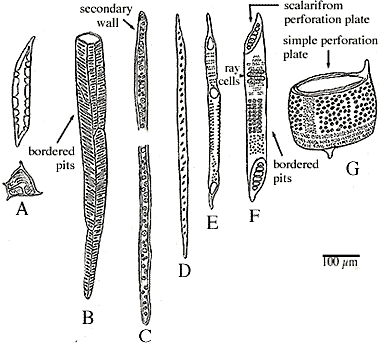
Xylem cells are
dead and empty of cell contents at maturity and essentially form tubes for
water transport. However, plants have no pumps to move water through these
hollow tubes. Thus water molecules are pulled in long, hydrogen-bonded chains
from rhizome to leaf. If the chain breaks, for example if a bubble forms in a
xylem cell, the involved cells lose their function and cannot be repaired.
Since xylem can be modeled as physical pipes following hydrodynamic principles,
the water-transport ability of ancient plants can be easily calculated.
Parenchyma cells are often present in xylem tissue, where they help maintain
water balance and carry out metabolism within the tissue. Because more than one
cell type is present in xylem, it is called a complex
tissue.
|
|
|
Figure 1.2: Ornamentation in xylem
as viewed in (A) transverse and (B) longitudinal section. Note annular,
spiral, scalariform and pitted sculpture. |
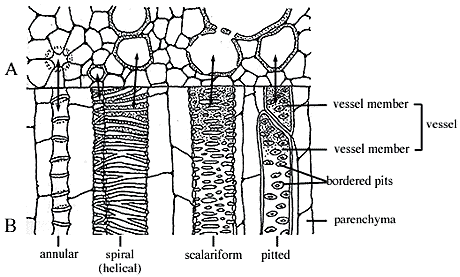
PHLOEM
Phloem
tissue transports photosynthetic products, other organic molecules (e.g., plant
hormones and waste products), and soluble nutrients throughout the plant.
Unlike xylem, phloem is alive at maturity, but usually with a much reduced cell
contents and no nucleus. This is logical because movement of material through
phloem tissue relies on solute gradients and some active transport that require
the activity of living cells. In non-angiosperm seed plants phloem elements
consist mostly of sieve cells
(Figure 1.3), while angiosperms have sieve tube
cells in association with parenchymatous companion
cells. Phloem fibers also provide some mechanical support. Phloem cells are
commonly unlignified so they do not preserve as
readily as xylem.
|
|
|
Figure 1.3: Phloem cell types. (A)
Longitudinal view of sieve-tube member and (B) sieve plate. (C-D) Sclerid reinforced with lignin. |

Some living land
plants, namely mosses, do not contain xylem and phloem. Instead, the gametophytes
of many mosses contain water conducting cells known as hydroids.
Like tracheids, hydroids are elongated cells with
oblique end walls, however they lack secondary ornamentation characteristic of tracheids. (Keep this fact in mind when we return to early
vascular plants in a few weeks.) Also like xylem, hydroids lack cell contents
at maturity and so appear empty. Some mosses also have solute-conducting leptoids surrounding a central bundle of hydroids. Leptoids are elongate cells that have nuclei and living
protoplasts and thus closely resemble the most generalized phloem cells of some
vascular plants. Hydroids may also be found in moss sporophytes, but leptoids
have been found only in the sporophytes of a few
genera.
|
|
|
Figure 1.4: Details of
conducting-tube cell wall construction. Modified from Kenrick
and Crane (1997). (A) S-type cells typical of some rhyniophytes,
(B) G-type cells typical of early lycopsids and zosterophylls, (C) P-type cells characteristics of Psilophyton
and many common living plants. |

The fossil record
of early land plants preserves a variety of other conducting-tube forms. Some
tubes are smooth and lack ornamentation. Others have helical thickenings with a
double-layer design in which a thin decay-resistant layer projects into the
cell lumen and a "spongy" outer layer extends outside the cell. This
S-type cell (Figure 1.4) is typical of early land plants like Rhynia.
G-type cells have ring-like or reticulate thickenings in which the inner layer
is decay-resistant and the outer layer is mineralized (organic material had
been replaced) in most fossils. This conducting cell type is typical of zosterophylls and early lycopsids.
The P-type cell has scalariform pitting typical of
most modern vascular plants.
Interpreting
Evolutionary Relationships
The term homology
was first introduced by zoologist Sir Richard Owen in 1843. The word is
derived from "homologia" in Greek which
means "agreement". Homology refers to structures or organs that have
evolutionary correspondence, regardless of their current function. The
homology of structure is based on similarities in morphology or developmental
origin. The wings of birds, forelimbs of a reptile and human arms are
homologous structures because they are all derived from the same primitive
structure in the common ancestor of these groups. On the other hand, analogous
structures may perform the same function, but are not derived from the same
structure in a common ancestor. The wings of bats and insects are therefore
analogous because they both function for flight, but are derived from
different primitive structures. Deciding whether
structures are homologous or analogous is key to
interpreting evolutionary relationships among organisms. However, making this
interpretation is seldom straightforward. For example, Johann Wolfgang von
Goethe noted in Metamorphosis
in Plants (1790) that plant organs such as cotyledons,
foliage leaves, bracts, and some flower parts are variously modified leaves.
Thus, these structures are homologous and we can begin to think about the
transformations necessary to develop their varied forms and new functions. We
might also consider the homology among conducting tubes in land plants. There
is certainly a variety of form. This could be evolutionary elaboration of a
single ancestral type (homology) or similar solutions to the problem of water
conduction that arose independently in several lineages (analogy). |
The
Organism- Building a Plant
|
|
|
Figure 1.5: The organ anatomy and
general vascular structure of a fern shoot. Note that the megaphyll
(= leaf or frond) has abaxial spore-producing structures. Use the
three-dimensional diagram to understand how a leaf gap relates to the
vascular cylinder of a siphonostele. |

The fern shoot in
Figure 1.5 will help you assemble all of the various tissue types into a plant.
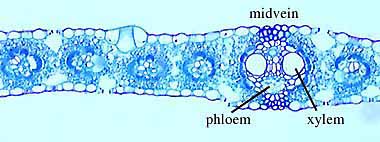
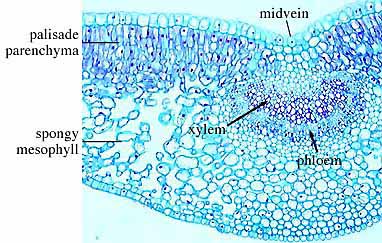
Examine sectioned stems of modern plants (Helianthus
- sunflower (VG 1:5), Pelargonium (VG 1:6), Ranunculus - buttercup (VG 1:7) and Lycopodium
(VG 1:9)) to identify
parenchyma, xylem, phloem, and other cell types if present. Note that they are
arranged together into distinct tissues. If you have never looked at the
cellular structure of a plant before, all of the cells may look alike to youÉdon't panic. In modern plant material, the cell types
are differentially stained and this will help you distinguish them at first
glance. However, don't rely on this crutch because the fossils don't come
stained. When you have sorted out which cell type is which, start to notice
features of the different cell types that would help you distinguish them in an
unstained preparation. For example, phloem cells tend to be a little bit
polygonal in contrast to the very round or oval xylem cells. A little time and
careful observation (aided by drawing) will train your eye quickly.
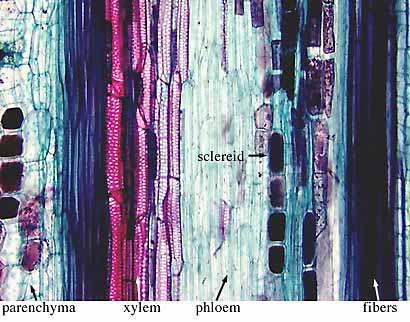
In slides of
macerated wood (wood that has been degraded by chemical treatment) you can observe
tracheids in three dimensions (VG 1:8). In thin sections of woody stems, note tracheids,
fibers, ray cells, and vessel elements (VG 2:9). To
find all of these characteristics, observe all three section planes: radial,
tangential, and transverse (Figure 1.6) (VG 2:10)(VG 2:11)(VG 2:12). Transverse
sections are taken perpendicular to the long axis of the stem (cross-section).
Radial sections are taken parallel to the long axis of the stem and cut through
the very center of the stem (on radii). Tangential sections are also taken
parallel to the long axis of the stem but are cut off center (along a tangent).
Each of these views allows you to see the rays in a different orientation. How
can you distinguish angiosperm wood from that of conifers and other plants?
|
|
|
Figure 1.6: Orientation of sections
for the study of wood anatomy. (A) Transverse; (B) longitudinal; (C)
tangential. |
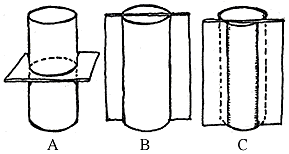
|
Note: Detailed, labeled
drawings will be valuable later as you try to recognize these various tissue
types in ancient plants. When making drawings, you are trying to compromise
between working quickly (so that you can get through all lab material) and
providing enough detail to later jog your memory. For example, when drawing a
stem cross section, it wouldn't be wise to try to draw every cell. Rather,
outline and label the general tissue types (e.g., vascular bundle, ground
tissue, cortex), then select one vascular bundle to draw in cellular detail,
labeling phloem, xylem, collenchyma, and ground
tissue. Artistic merit is not important, utility is. Make sure that your
drawings include features important for recognizing the structure or taxon. Also, label your drawings clearly so that anyone
(even you when you study for the exam) can interpret them. |
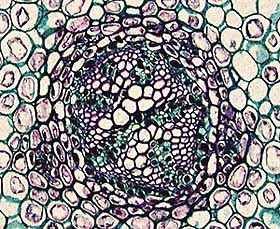
The
Stele
The plant stele
consists of the primary vascular system of the plant axis (stem) and its
associated ground tissues (e.g., pith). The stele consists solely of primary
tissues differentiated from procambial strands derived from the apical meristem. Secondary
vascular tissue (wood consists of secondary xylem) is derived from a vascular
cambium.
|
|
Figure 1.7: Diagram of a plant shoot
showing apical meristem, the center of primary
growth, a node with leaves and branch bud, and the internode
region between nodes. |

Because phloem is
rarely preserved on fossils (because the cell walls are not reinforced with
lignin, these cells are often crushed or destroyed chemically during
preservation), it is the structure of the xylem --particularly primary xylem--
that is of particular interest for paleobotany. Understanding
stele types is necessary for interpreting vascular system evolution and for
identifying plant axes. The most comprehensive review of stelar
morphology in living and fossil plants is that of Beck, Schmid
and Rothwell (1982; Botanical Review 48(4):691-817) and a companion
paper by Schmid (pp. 817-931) in the same volume. You
will note that Rudolf Schmid is a professor in
Integrative Biology; he teaches the popular California Plant Life course.
The array of
descriptive stele types is overwhelming but don't panic, you don't need to
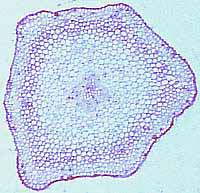
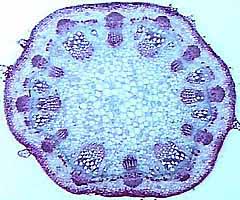
memorize them all. You should become familiar with the protostele (Figure 1.8)(VG 2:1)), a solid
interior core of xylem surrounded by a cylinder of phloem, siphonostele (VG 2:3), a central pith
(parenchyma) surrounded by a cylinder of vascular tissue, and eustele(VG 2:4)(VG 2:5), separate vascular
bundles in the cortex
with phloem to the outside of the xylem. This stele is characteristic of dicot angiosperms. You will also encounter dictyosteles, which are complex siphonosteles
in which the vascular cylinder is broken up by many leaf gaps.
This gives the stem cross section the appearance of concentric, broken rings.
Similarly, actinosteles (VG 2:2) are protosteles in which the central vascular strand is lobed, givng is a star-shaped silouette in cross section. There are many other
elaborations on these basic steles that will crop up occasionally--be on the
lookout!
|
|
|
Figure 1.8: The basic stele types in
vascular plants. (A) Protostele, (B) siphonostele, (C-D)
eustele. |

One useful
character of stelar development is the maturation of
the primary xylem. The earliest maturing xylem cells are called protoxylem. These xylem elements are generally small
and narrow. Later maturing and larger elements are known as metaxylem. If the protoxylem
strands are external to the metaxylem, the stele is exarch (VG 2:6); if protoxylem is internal to the metaxylem,
the stele is endarch (VG 2:7); if metaxylem surrounds the protoxylem,
the stele is mesarch (VG 2:8).
Combining patterns
of xylem maturation with the relative position of phloem and xylem permits a
very precise description of the stele in a very few words. For example the
sunflower stem (Helianthus)
possesses an endarch ectophloic (phloem on the outside) eustele.
Do you believe me? Don't look at the slide, check your drawing!
|
|
|
Figure 1.9: Diagrammatic
representation of the relationship between stele, leaf trace, and leaf gap in
three dimensions. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Leaf gaps
are features often found in siphonosteles (Figure
1.9). They are discontinuities in the vascular cylinder that occur where leaf traces
(the vascular bundles supplying leaves) depart from the stele. If you were to
examine serial sections up through the plant axis, leaf gaps would originate
and close all along the stele as leaf traces arose at each node.
Leaf gaps occur
only in siphonosteles and related types. In protosteles, leaf traces simply diverge from the solid
vascular cylinder. The areas between the vascular bundles in a eustele are not leaf gaps. In eusteles,
leaf traces arise from individual vascular bundles as if they were tiny protosteles.
In sections of
woody stems, you may note growth rings in the secondary xylem
. In seasonal climates, growth varies throughout the year producing
annual rings (VG 2:9)(VG 2:10). The diameter
of the ring and the size of the cells can tell much about conditions within and
between growing seasons. Distinguish primary and secondary tissues in these
stems, and note the position of the vascular
cambium, although you won't be able to actually see it. Note rays (parenchyma
cells revisited) in woody tissue. Diagram (don't draw in cellular detail) the periderm, the protective tissue outside of the wood. Periderm is composed of cork, a
secondary tissue derived from activity of the cork
cambium.
This is far from
an exhaustive review of tracheophyte anatomy and morphology. Undoubtedly many
more anatomical terms will come up as our survey of fossil plants continues and
as you read the primary literature. Feel free to refer to the several
references mentioned here or ask when something seems unclear.
Now, return to the
coal ball sections; can you identify plant tissues in the coal balls? What are
they? What stele types are present?
Questions for
Further Thought
These questions
encourage you to think in more detail about topics covered in this lab. Becaure they are thought questions, there aren't
necessarily single "right" answers. Don't be surprised if some of
these questions reappear on a quiz or midterm.
1.
What distinguishes a root from a rhizome? From an evolutionary
perspective, why might this distinction be important?
2.
How would you define a "tree"? Tuck your definition
away for a few weeks until we study the arborescent lycopsids of the Paleozoic-are they "trees" or
overgrown "herbs"?
3.
"Cavitation" is the
process of bubbles forming in xylem. When such bubbles form (say by an ice
crystal shrinking as it thaws to liquid), they break the continuous chain of
water molecules from root to leaf and render the xylem non-functional. What
sort of ecological implications might this phenomenon have for the plant?
4.
Are the conducting tubes in tracheophytes
and bryophytes homologous? What additional information would be needed to
increase confidence in your answer?